
Abstract
Thymosin Beta-4 (T$beta$4), and its synthetically derived bioactive fraction TB-500, are low molecular weight, naturally occurring proteins expressed ubiquitously across numerous tissue environments. Initially characterized for their pivotal roles in cardiovascular repair and wound healing paradigms, recent advanced in vitro and preclinical modeling has pivoted toward exploring their potent regenerative capacities within the complex microenvironment of the hair follicle. The hair cycle is a tightly regulated orchestration involving stem cell quiescence and activation, vascular remodeling, and robust extracellular matrix (ECM) reorganization. This expansive technical brief deconstructs the biophysical and biomolecular mechanisms by which T$beta$4/TB-500 governs critical pathways such as actin cytoskeletal dynamics, Wnt/$beta$-catenin signaling, pro-angiogenic VEGF upregulation, and matrix metalloproteinase (MMP) secretion within isolated cellular models.
1. Introduction: The Hair Follicle as a Complex Mini-Organ
The hair follicle represents uniquely dynamic, self-renewing tissue characterized by distinct cyclical phases: anagen (active growth), catagen (regression and apoptosis), and telogen (relative rest). The transition from telogen back into anagen requires the highly coordinated activation of multipotent stem cells residing within the specialized anatomical niche known as the “bulge,” located at the insertion site of the arrector pili muscle.
Initiation of a new cycle depends on cross-talk between these bulge stem cells and the underlying mesenchymal dermal papilla (DP) cells. In vitro studies focusing on these specific cellular populations frequently require exogenous mitogenic factors to maintain viability and observe differentiation pathways. Structurally, Thymosin Beta-4 is widely recognized as a major actin-sequestering protein in eukaryotic cells. By investigating its synthetic analogue, TB-500, researchers possess a stable, structurally precise peptide segment perfect for isolating these intricate biochemical pathways in controlled laboratory environments.
2. Cytoskeletal Dynamics and Actin Sequestration
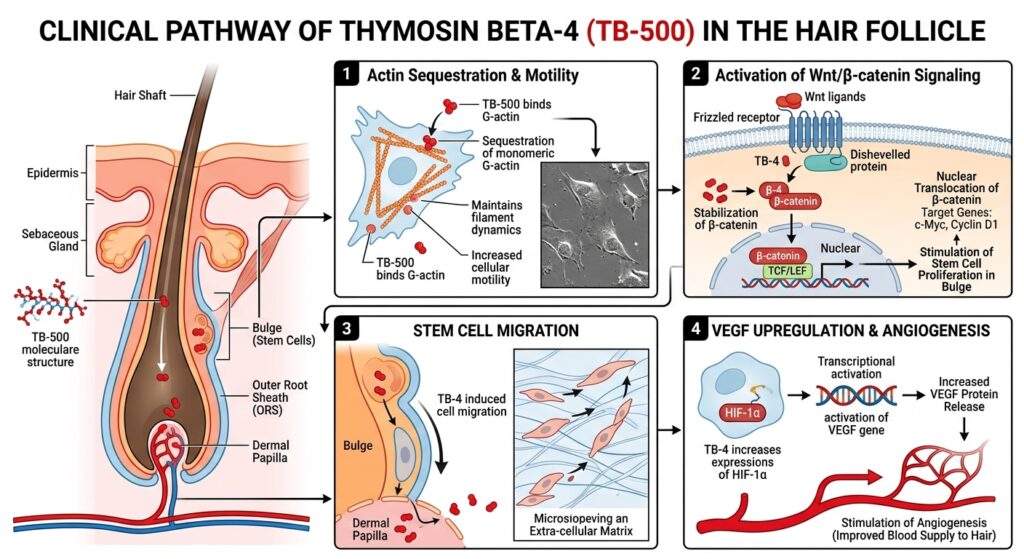
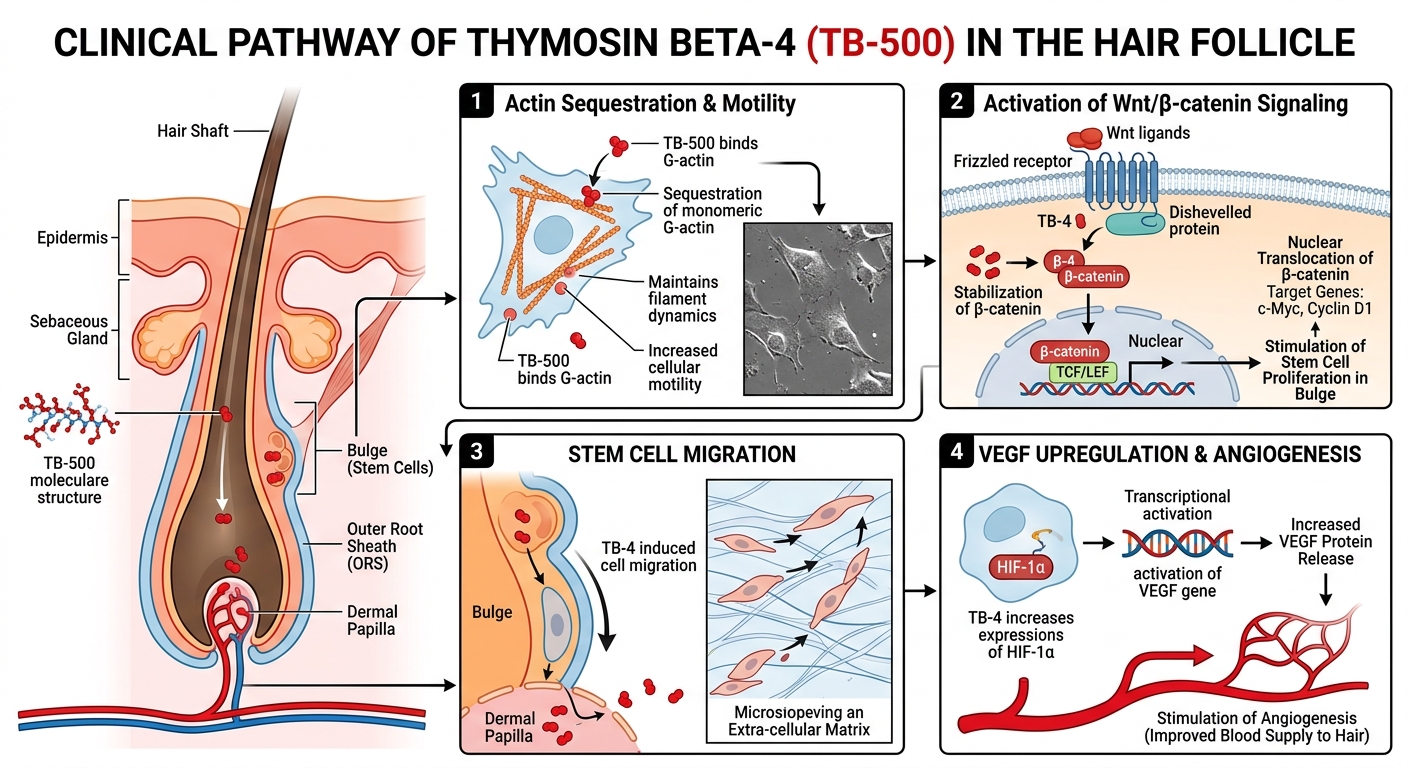
The most fundamental, thoroughly documented mechanism of T$beta$4 across all cell types is its interaction with actin. The cellular cytoskeleton is highly dependent on continuous polymerization (forming fibrous F-actin) and depolymerization (returning to monomeric G-actin). This dynamic flux is the engine driving cell crawling, migration, and structural morphology.
T$beta$4 binds specifically to G-actin monomers in a 1:1 complex. By sequestering G-actin, it inhibits its spontaneous polymerization into F-actin. While counterintuitive, maintaining a readily available pool of unpolymerized G-actin is crucial. When extracellular signals trigger directed migration—such as those required for bulge stem cells to migrate downward to reform the hair bulb during early anagen—T$beta$4 releases G-actin rapidly in response to localized spatial cues.
In vitro tracking assays utilizing fluorescently labeled follicular keratinocytes and stem cells exposed to TB-500 demonstrate significantly accelerated rates of cell spreading and directional migration compared to control models. The managed, localized release of G-actin enables the rapid filament turnover required at the leading edge (lamellipodia) of migrating cells originating from the follicular bulge.
3. Activation of Follicular Stem Cell Niche
The transition out of the dormant telogen phase requires breaking stem cell quiescence. In vitro models utilizing isolated hair follicle bulge stem cells provide remarkable insights into T$beta$4’s regulatory capabilities over stem cell fate.
3.1 Orchestration of the Wnt/$beta$-Catenin Pathway
The Wnt signaling cascade is the master regulator orchestrating hair follicle morphogenesis and the initiation of the anagen phase. When Wnt ligands bind to their respective Frizzled receptors on the cell surface, they shut down the degradation complex that normally destroys $beta$-catenin. Consequently, $beta$-catenin stabilizes, accumulates in the cytoplasm, and translocates into the nucleus. Here, it acts as a transcriptional co-activator for genes that drive cell proliferation and lineage-specific differentiation.
In vitro investigations reveal that administration of T$beta$4 analogues directly activates the canonical Wnt pathway within follicular matrices. Co-culture assays of dermal papilla and bulge cells demonstrate that the presence of T$beta$4 elevates nuclear accumulation of $beta$-catenin, confirming a direct upstream stimulatory effect that mimics the natural signaling required to spark new follicular growth cycles.
3.2 Proliferation of Outer Root Sheath (ORS) Cells
The outer root sheath is critical for anchoring the hair shaft and serving as a conduit for migrating progenitors. Proliferation assays utilizing tritiated thymidine or BrdU incorporation in isolated ORS keratinocytes show a marked, dose-dependent increase in mitotic activity when cultured in media supplemented with T$beta$4/TB-500 analogues. This robust cellular expansion is a requisite step for constructing the physical infrastructure of a new, actively growing hair shaft.
4. Angiogenesis and Extracellular Matrix (ECM) Remodeling
The metabolic demands of an actively dividing hair matrix during anagen are tremendous. Consequently, robust vascularization around the growing follicle is absolutely essential for nutrient delivery and the removal of metabolic waste.
4.1 Up-Regulation of Vascular Endothelial Growth Factor (VEGF)
T$beta$4 operates as a potent pro-angiogenic factor. In vitro cultures utilizing isolated dermal papilla cells point to a powerful secondary mechanism. Upon exposure to T$beta$4, these specialized fibroblasts significantly upregulate the expression and secretion of VEGF, often demonstrating a several-hundred-percent increase relative to baseline controls.
VEGF subsequently acts on neighboring endothelial cells to promote the sprouting and formation of new capillary networks. This localized angiogenic drive ensures that the developing hair bulb is suffused with adequate blood supply, effectively preventing premature entry into the catagen phase driven by nutrient starvation or hypoxia.
4.2 Modulation of Matrix Metalloproteinases (MMPs)
Cells do not exist in a vacuum; they are encased in a dense, restrictive extracellular matrix (ECM). For cells to migrate—whether they are stem cells exiting the bulge or endothelial cells forming new vessels—they must digest their way through this barrier.
T$beta$4/TB-500 influences this process by stimulating the targeted secretion of specific matrix metalloproteinases, most notably MMP-2. In scratch-wound assays and 3D transwell migration models, elevated MMP-2 activity mediated by T$beta$4 allows for transient, controlled degradation of collagen and elastin fibers within the ECM. This targeted structural loosening is a necessary biophysical requirement, enabling the rapid geometric reorganization and cellular influx characteristic of early anagen onset.
5. Modulation of Inflammatory Cytokines in the Dermal Milieu
Micro-inflammation within the dermal layers is widely suspected to contribute to the miniaturization of hair follicles, leading to their progressive dysfunction. Elevated levels of pro-inflammatory cytokines, specifically Tumor Necrosis Factor-alpha (TNF-$alpha$) and Interleukin-1 beta (IL-1$beta$), are potent negative regulators of the hair cycle.
In vitro cell culture models incorporating immune cells (like macrophages) or stressed dermal fibroblasts demonstrate that the introduction of T$beta$4 significantly suppresses the transcription and subsequent release of these inflammatory markers. By attenuating the localized inflammatory cascade, T$beta$4 effectively shields the vulnerable follicular stem cells and DP cells from apoptotic signals, fostering a biochemically pro-survival environment permissive to sustained cellular division and differentiation.
6. Conclusion
The synthesis of highly pure TB-500 continues to afford researchers a pristine tool to meticulously map the complex intracellular networks governing tissue regeneration. Within the context of hair follicle biology, in vitro evidence consistently outlines a sophisticated, multi-pronged mechanism of action. By intimately regulating G-actin sequestration for cellular migration, activating the paramount Wnt/$beta$-catenin master regulatory pathway, driving robust localized angiogenesis via VEGF, and remodeling the physical barriers of the ECM via MMP-2, Thymosin Beta-4 represents a dominant modulatory polypeptide in follicular biochemistry.
Scientific References & Further Reading:
- Philp, D., et al. (2004). Thymosin beta-4 induces hair growth in mouse models via stem cell activation. PNAS, 101(30), 10803–10808.
- Zhuang, Y., et al. (2021). Thymosin $beta$4 Promotes Hair Growth via Wnt/$beta$-Catenin Pathway. Frontiers in Pharmacology, 12, 608466.
- Meier, N., et al. (2007). Thymosin beta4 promotes hair growth. Journal of Investigative Dermatology, 127(12), 2743–2744.
- Crockford, D. (2007). Development of thymosin beta4 for treatment of cardiovascular diseases. Expert Opinion on Biological Therapy, 7(3), 335–347.
- Cha, H. J., et al. (2010). Thymosin beta 4 induces hair growth via stem cell migration and Wnt signaling pathway. Annals of Dermatology, 22(3), 324–328.
(Disclaimer: The content detailed above is intended strictly for in vitro laboratory research and academic reference. Synthetic peptides discussed herein are not approved, designed, or strictly evaluated for human consumption, diagnostics, or therapeutic interventions.)