
Abstract
Semax is a synthetic heptapeptide (Met-Glu-His-Phe-Pro-Gly-Pro) modeled after a biologically active fragment of Adrenocorticotropic Hormone (ACTH 4-7). Originally designed to isolate the cognitive-enhancing properties of ACTH devoid of steroidal axis stimulation, Semax exerts profound modulatory effects on critical neurotrophic factors including Brain-Derived Neurotrophic Factor (BDNF) and Nerve Growth Factor (NGF). However, like all naked native peptides, Semax suffers from extreme vulnerability to enzymatic cleavage when placed in raw physiological media during in vitro testing. To circumvent this, biochemists developed N-Acetyl Semax Amidate (Ac-Semax-$NH_2$). This comprehensive structural analysis delineates how specific terminal molecular capping strategies drastically alter the proteolytic degradation curve of the peptide, while preserving its potent intracellular signaling capacity regarding the CREB-dependent upregulation of essential neuroplasticity proteins.
1. Introduction: The Neurotrophic Directive of the ACTH Fragment
Neurogenesis and synaptic plasticity—the mechanisms underlying learning, memory, and cognitive repair—are heavily dependent on the continuous biosynthesis of neurotrophins. BDNF and NGF are paramount in this equation, promoting the survival of existing neurons and directing the growth and differentiation of new synapses.
The endogenous melanocortin system, specifically fragments of ACTH, has long been known to influence learning and memory. Semax isolates this specific neuro-active fragment (ACTH 4-7: Met-Glu-His-Phe) and stabilizes it by appending a C-terminal tripeptide domain (Pro-Gly-Pro), which naturally resists certain endopeptidases. In controlled in vitro models utilizing isolated hippocampal and basal forebrain neuronal cultures, the introduction of Semax reliably triggers a rapid, marked upregulation in the mRNA transcription and subsequent translation of both BDNF and NGF.
2. Intracellular Mechanisms: The CREB Pathway
While the exact primary receptor for Semax remains a subject of ongoing investigation (with strong evidence pointing toward melanocortin receptors MC4R and MC5R), the intracellular cascade it triggers is well documented in cellular assays.
Upon binding to the neuronal surface, Semax activates secondary messenger systems that ultimately converge on the activation of several kinase cascades (frequently involving the cAMP/PKA or MAPK/ERK pathways). These kinases translocate to the nucleus where they heavily phosphorylate the cAMP Response Element-Binding protein (CREB).
Activated, phosphorylated CREB serves as the master transcriptional switch for neuroplasticity. It binds directly to the promoter regions of the BDNF and NGF genes, aggressively driving their transcription. In vitro analyses confirm that exposure to Semax yields highly elevated levels of BDNF protein in the culture media shortly after administration, confirming its potent neurogenic drive. Additionally, it exerts significant neuroprotective effects by modulating glutamate receptor expression, thereby buffering neurons against excitotoxic damage commonly seen in ischemic cellular models.
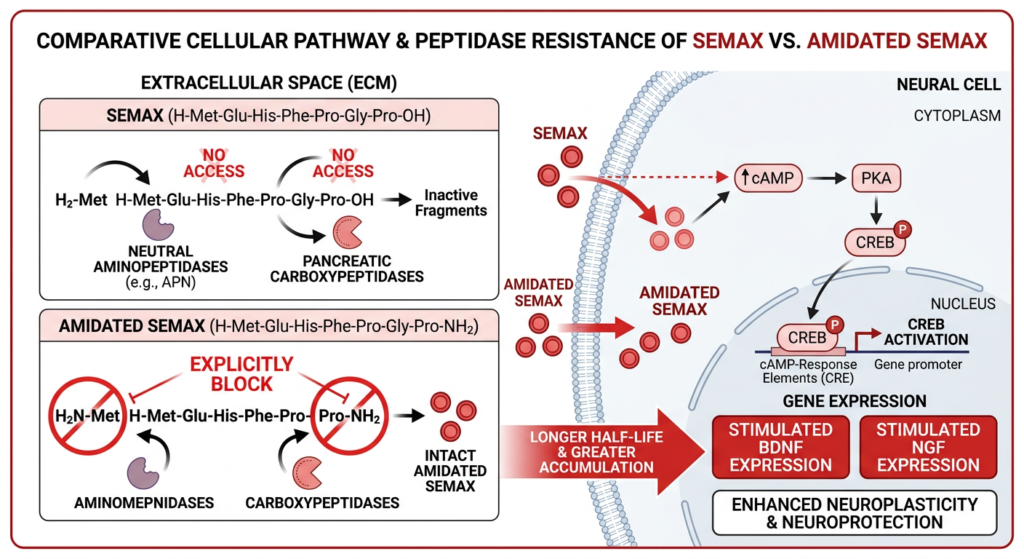
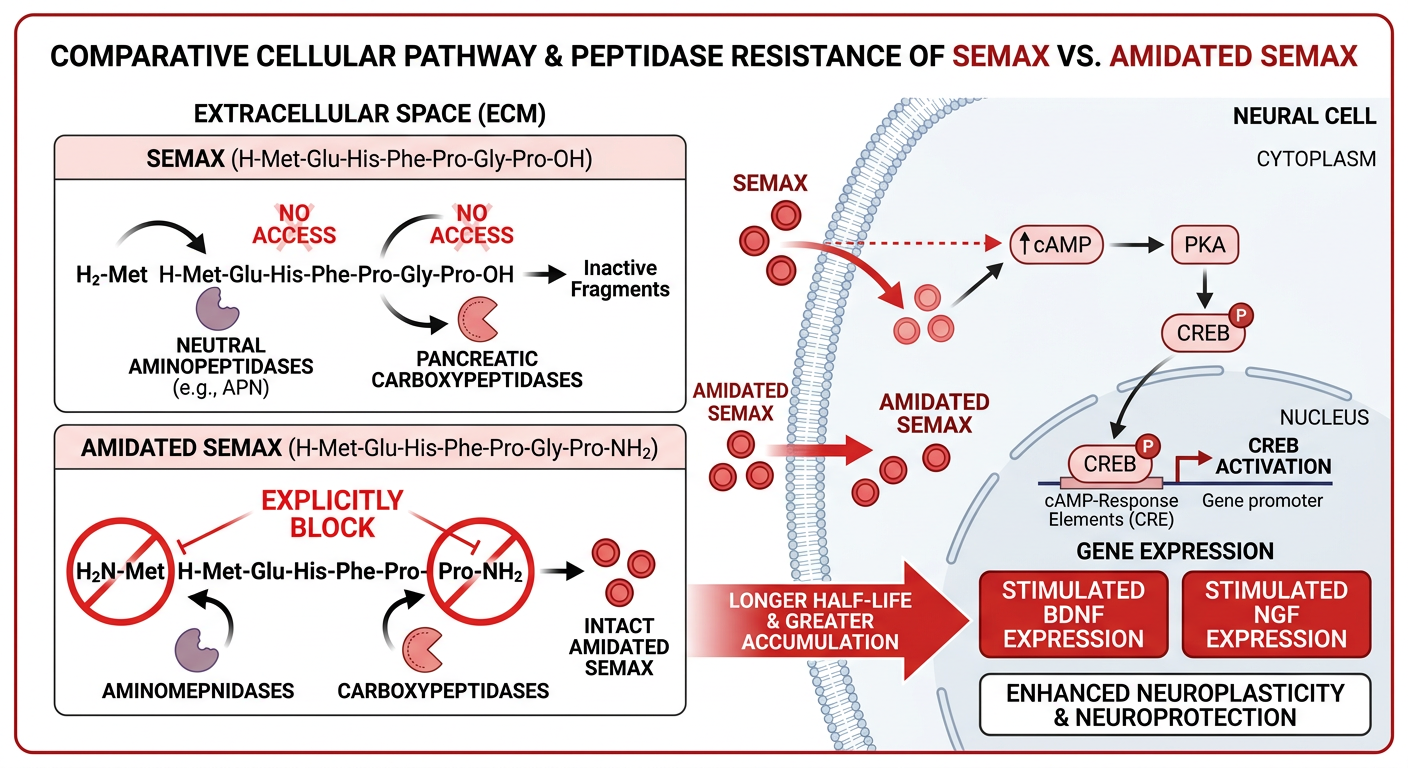
3. The Vulnerability of the Naked Peptide: Proteolytic Degradation
The primary obstacle encountered when utilizing standard Semax in vitro—particularly when the culture medium contains serum or complex tissue homogenates—is its extreme susceptibility to enzymatic degradation. The peptide bonds holding the sequence together are prime targets for aggressive physiological proteases.
Standard Semax possesses a “free” or naked N-terminus (an unbound amine group on the Methionine residue) and a naked C-terminus (an unbound carboxyl group on the terminal Proline).
- Aminopeptidases: Encrusted on cellular membranes and floating in serum, these enzymes act like chemical pac-men, recognizing the naked N-terminus and sequentially cleaving off amino acids one by one from the “front” of the peptide.
- Carboxypeptidases: Conversely, these enzymes recognize the naked C-terminus and aggressively chew away at the peptide from the “back.”
This dual enzymatic assault drastically reduces the half-life of naked Semax in complex physiological media, often reducing it to mere minutes. This rapid degradation severely limits the peptide’s “area under the curve” (AUC) and restricts the duration of its neurotrophic signaling.
4. The Engineering of N-Acetyl Semax Amidate
To engineer a more stable molecule for prolonged in vitro study and eventual pharmacokinetic application, chemists employ terminal capping strategies to physically block the vulnerable ends of the peptide. N-Acetyl Semax Amidate represents the pinnacle of this structural fortification.
4.1 N-Terminal Acetylation
The naked amine group on the N-terminal Methionine residue is chemically bound to an acetyl group ($CH_3CO-$). This seemingly minor modification fundamentally alters the chemical signature of the peptide’s front end. By removing the positive charge and altering the steric bulk, the N-terminus is rendered effectively “invisible” to aminopeptidases. They can no longer recognize or bind to the peptide, completely shutting down N-to-C directional degradation. Furthermore, acetylation reduces the affinity of disruptive metal ions (like $Cu^{2+}$) to the peptide, further enhancing its stability in solution.
4.2 C-Terminal Amidation
Simultaneously, the naked carboxyl group on the C-terminal Proline is replaced with an amide group ($-NH_2$). This modification effectively neutralizes the negative charge at the C-terminus. By capping this end, carboxypeptidases are sterically and chemically blocked from initiating degradation from the rear. Amidation also mimics the natural post-translational modifications of many endogenous neuropeptides, often enhancing receptor binding affinity.
5. Comparative Stability in Vitro
By utilizing both an N-terminal “shield” (acetylation) and a C-terminal “shield” (amidation), N-Acetyl Semax Amidate forces degradative enzymes to rely solely on endopeptidases (which attempt to cut the peptide in the middle). However, the internal sequence of Semax, guarded by the bulky Pro-Gly-Pro tail, is already somewhat resistant to endopeptidase cleavage.
When subjected to rigorous in vitro stability assays using human or rat plasma equivalents, the modified Ac-Semax-$NH_2$ demonstrates a vastly superior survival curve. While the naked Semax may be degraded within minutes, the acetylated and amidated version maintains its structural integrity exponentially longer, often persisting for hours.
Crucially, because the core active sequence (the ACTH 4-7 fragment) remains structurally unchanged, Ac-Semax-$NH_2$ retains the full neurotrophic potency of the original molecule. It activates the exact same CREB pathways and drives the identical synthesis of BDNF and NGF, but it remains viable in the culture dish for significantly longer, leading to a much more profound and sustained biological effect.
6. Conclusion
The biochemical evolution from standard Semax to N-Acetyl Semax Amidate perfectly illustrates the necessity of structural optimization in peptide science. While both compounds powerfully orchestrate the upregulation of vital neuroplasticity proteins (BDNF/NGF) via CREB phosphorylation, the naked peptide is severely handicapped by its rapid destruction via amino- and carboxypeptidases. By strategically capping the molecular termini through acetylation and amidation, researchers have engineered a highly stable, enzymatically resistant analogue. This allows for prolonged receptor engagement and expanded utility in rigorous in vitro models studying advanced neuroprotection and enhanced cognitive architecture.
Scientific References & Further Reading:
- Magrì, A., et al. (2016). Influence of the N-terminus acetylation of Semax (ACTH(4–10) analog) on copper(II) and zinc(II) coordination and biological properties. Journal of Inorganic Biochemistry.
- Potaman, V.N., et al. (1991). N-terminal degradation of ACTH(4–10) and its synthetic analog Semax by rat blood enzymes. Biochemical and Biophysical Research Communications.
- Eremin, K. O., et al. (2005). Semax, an ACTH(4-10) analogue with nootropic properties, activates dopaminergic and serotonergic brain systems in rodents. Neurochemical Research, 30(12), 1493-1500.
- Agapova, O. A., et al. (2008). The effect of the synthetic peptide Semax on the expression of neurotrophic factors in the rat brain. Neuroscience and Behavioral Physiology, 38(3), 285-288.
(Disclaimer: The content detailed above is intended strictly for in vitro laboratory research and academic reference. Synthetic peptides discussed herein are not approved, designed, or strictly evaluated for human consumption, diagnostics, or therapeutic interventions.)